Last Modified: April 9, 2020
Instruments: unknown, Inductively Coupled Plasma Optical Emission Spectrometer, X-ray diffractometer
| URL | https://www.bco-dmo.org/dataset/806957 |
|---|---|
| Download URL | https://www.bco-dmo.org/dataset/806957/data/download |
| Media Type | text/tab-separated-values |
| Created | March 25, 2020 |
| Modified | April 9, 2020 |
| State | Final no updates expected |
| Brief Description | Table 2 |
X-ray diffraction (XRD)
Powdered samples were examined for mineral presence using a Rigaku Ultima III X-ray Diffraction (XRD) System at the Institute of Imaging and Analytical Technologies (i2AT) at Mississippi State University. These analyses were conducted at 40 kV and 40 mA.
Elemental analysis of experimental aragonite crystals and experimental fluids with ICP-MS
Crystals were dissolved and solutions were analyzed by ICP-MS. Experimental fluids were also analyzed with ICP-MS, as well. Analyses were conducted on an Agilent 7900 quadrupole ICP-MS equipped with a glass nebulizer at the University of Rochester (Rochester, NY). Prior to analysis, fluid samples were diluted with 2% trace metal grade HNO3 (Fisher) in the volume proportion of 1:100. Aragonite crystals were dissolved in HNO3 in the mass proportion of 1:1000. Experimental fluid samples were analyzed using a 5-point calibration curve (one blank and four standards). Seawater standards were prepared by diluting a seawater certificate reference material (CRM-SW; High-Purity Standards) to a final concentration of ~300, 200, 150, and 80 ppm. Experimental crystals were analyzed using a 5-point calibration curve (one blank and four standards). Calcium carbonate standards were prepared by dissolving and diluting the original MACS-3 powder to a final concentration of ~111, 55, 27, and 11 ppm. Because S concentration was not reported in the certificate of analysis provided by the USGS, we added a small amount of sulfur single element standard (Inorganic Ventures) to a final concentration of ~1.5, 0.75, 0.375, and 0.150 ppm, in each MACS-3 standard we prepared. Samples, blanks, and standards were also spiked with 2 ppb indium, which was used as the internal standard to check for possible instrumental drift during each analytical session.
The instrument was tuned at the beginning of each analytical session (two analytical sessions in two consecutive days). 24Mg, 34S, 43Ca, 88Sr, 138Ba, and 238U were analyzed. Integration times were the following: 0.3 s for 24Mg; 0.12 s for 43Ca, 88Sr, and 138Ba; 0.99 s for 238U. The calibration curve was run before analyzing each set of samples. For each element analyzed, the correlation among standards was 0.9923 or better. Elements were identified using three peaks. Each analysis included three replicates, with 100 sweeps/replicate. The carrier gas (argon) flow was set at 1.15 L/min. In addition, we used helium flow of 4.2 milliliters per minute in the collision reaction cell in the ICP-MS to minimize oxide interference. At the end of each tuning, oxides and doubly-charged ion interferences were below 0.8% and 1.4%, respectively. Data were computed automatically during the run using the Agilent Mass Hunter 4.1 workstation software v. C.01.01.
Results
XRD spectra identified the presence of aragonite and monohydrocalcite (Table 2). Only aragonite crystallized at intermediate pressure (110 bars). Calculations of fluid carbonate chemistry were conducted with a CO2SYS spreadsheet (Lewis and Wallace, 1998) and presented in Table 1. There, measured pH and total alkalinity (TA) were used to calculate the concentration of CO32-, which together with Ca2+ is necessary to calculate fluid saturation states with respect to aragonite (Omega-Ar). Precipitation started at saturation state, which exceeds that of artificial seawater (ASW) by the factor of ~25. Over the course of the experiments, Omega-Ar decreased back to 2, i.e. Omega-Ar value of ASW prior to the addition of Na2CO3. Table 2 contains E/Ca of experimental products (solids and liquids) as well as the Doener-Hoskins apparent partition coefficients between solid and fluid (KE). Fluid composition changed during individual experiments, and therefore, values of KE were calculated using the Doener-Hoskins relationship:
KE= log(1+ mE^aragonite /mE^fluid) / log(1+ mCa^aragonite /mCa^fluid) (1)
where m^aragonite is the total number of moles of element (i.e., Mg, S, Sr, Ba, or U) or Ca in the final precipitate, and m^fluid is the total number of moles of element or Ca in the final fluid (Doerner and Hoskins, 1925).
Software products used:
XRD: Jade and Microsoft Excel
ICP-MS: Agilent Mass Hunter 4.1 workstation software v. C.01.01 and Microsoft Excel
BCO-DMO Data Manager Processing Notes:
– added a conventional header with dataset name, PI name, version date
– modified parameter names to conform with BCO-DMO naming conventions
– blank values in this dataset are displayed as “nd” for “no data.” nd is the default missing data identifier in the BCO-DMO system.
– reformatted table so each column represent only one parameter
– removed quotes, replaced commas with semicolons
Custom designed pressure chamber used to simulate the high-pressure conditions at the seafloor for this set of experiments (designed at Rensselaer Polytech Institute, Department of Earth and Environmental Sciences).
No relevant match in BCO-DMO instrument vocabulary.
Also referred to as an Inductively coupled plasma atomic emission spectroscope (ICP-AES). These instruments pass nebulised samples into an inductively-coupled gas plasma (8-10000 K) where they are atomised and excited. The de-excitation optical emissions at characteristic wavelengths are spectroscopically analysed. It is often used in the detection of trace metals.
Located at at Mississippi State University
Instruments that identify crystalline solids by measuring the characteristic spaces between layers of atoms or molecules in a crystal.
The type of analysis: solids; fluids; or KE (Doener-Hoskins partition coefficients between solid and fluid)
water pressure at measurement; depth reported as pressure; positive number increasing with water depth
| Name | Affiliation | Contact |
|---|---|---|
| Rinat Gabitov | Mississippi State University (MSU) | ✓ |
| Karyn L. Rogers | Mississippi State University (MSU) | ✓ |
| Chiara Borrelli | University of Rochester | |
| Nancy Copley | University of Rochester | |
| Nancy Copley | Woods Hole Oceanographic Institution (WHOI BCO-DMO) |
| Project Title | Biologically induced methane oxidation and precipitation of carbonate minerals: An experimental study |
|---|---|
| Acronym | Deep Sea Geochem CaCO3 |
| URL | https://www.bco-dmo.org/project/743426 |
| Created | August 9, 2018 |
| Modified | September 14, 2018 |
Biologically-mediated CaCO3 precipitation widely occurs in aquatic systems and is often directly linked to the metabolic activity of microorganisms, which could significantly affect the local environment. An example is oxidation of methane and reduction of sulfate mediated by a consortium of Bacteria and Archaea. In order to investigate geochemistry of CaCO3precipitated abiotically and under microbial activity experimental work was initiated. The abiotic experiments were performed at different temperatures and pressures (nitrogen and nitrogen-methane mixture). Further geochemical analyses will allow evaluating of the effect of total pressure and methane partial pressure on the geochemistry of CaCO3. Sulfate reducing bacterial (Desulfovibrio salexigens) was successfully cultured and precipitation experiments on microbially mediated CaCO3 are in progress. This work is relevant to C-DEBI Research Theme I (Activity in the Deep Subseafloor Biosphere: function & rates of global biogeochemical processes) because carbonate growth rate is linked to microbial activity and the rate of methane oxidation.
| Name | Affiliation | Role |
|---|---|---|
| Rinat Gabitov | Mississippi State University (MSU) | Principal Investigator |
| Karyn L. Rogers | Rensselaer Polytechnic Institute (RPI) | Co-Principal Investigator |
| Chiara Borrelli | University of Rochester | Contact |
| URL | https://www.bco-dmo.org/dataset/806920 |
|---|---|
| Download URL | https://www.bco-dmo.org/dataset/806920/data/download |
| Media Type | text/tab-separated-values |
| Created | March 25, 2020 |
| Modified | April 8, 2020 |
| State | Final no updates expected |
| Brief Description | Table 1: Carbonate chemistry |
Crystallization experiment
In order to simulate the high-pressure conditions at the seafloor, three laboratory experiments involving aragonite precipitation were conducted. Two experiments were conducted using a high-pressure apparatus (Figure 1). The control experiment was performed at atmospheric pressure and 8°C in a container immersed in a constant temperature bath. The experimental fluid (growth medium) was prepared by mixing artificial seawater (ASW) and small aliquots of Na2CO3. The addition of Na2CO3 allowed the saturation state of the fluid with respect to aragonite to be increased, in order to promote nucleation and crystallization on carbonate minerals. Artificial seawater was prepared by dissolving 32 g of Instant Ocean salt (Instant Ocean Spectrum Brands) mix in 1 kg of distilled water and filtering it through a 0.2 µm Nalgene filter. A small amount of 0.1M Na2CO3 was then introduced into the continuously stirred artificial seawater using a syringe pump set at a rate of ~1 ml/min, to reach a final volume ratio of Na2CO3 to artificial seawater of ~1:13. During the addition of Na2CO3, the solution was in contact with the atmosphere. This procedure homogenized the fluid and also precluded aragonite nucleation prior to the beginning of the experiment. The absence of crystal nucleation was confirmed visually by monitoring the solution pH, which remained stable for the 1-2 hours needed for the experiment setup. The observation that aragonite crystallization did not start prior to the beginning of the experiment is consistent with previously published data, which show that 6 hours elapsed before onset of crystallization in the solution similar to the one we used (7.4 mmol of Na2CO3 :1 liter of seawater) (Pytkowicz, 1965).
Petrographic glass slides frosted with silicon carbide (SiC) abrasive paper were used to promote aragonite crystallization along scratches. For each experiment, one slide was placed into the high-pressure vessel prior to the transfer of the experimental fluid. The experimental fluid was then placed into the vessel after it was precooled to 8°C. After that, the air space above the solution (<200 cm3) was purged with nitrogen three times before the pressure was increased to a desired value (110 or 345 bars). The experimental fluid was sampled daily using a valve on the pressure vessel (Figure 1) and pH was measured to estimate time of nucleation and crystallization of CaCO3. For pH measurements, electrode calibration and measurements were conducted at temperatures similar to those of the experiment. NBS buffer solutions with pH of 4, 7, and 10 were used for electrode calibration. The pH initially decreased due to carbonate precipitation and reached near-constant values less than 170 hrs from the beginning of the experiments, suggesting that most of the CaCO3 precipitated in the first few days of each experiment. At the end of each run, the experimental products were extracted from the pressure vessel. Crystals (<10 µm in size) were rinsed with distilled water and dried, whereas the experimental fluid was filtered and refrigerated for subsequent elemental analyses. The alkalinity of the final fluid was measured the day after the end of each experiment. Experimental parameters are presented in Table -1.
BCO-DMO Data Manager Processing Notes:
– added a conventional header with dataset name, PI name, version date
– modified parameter names to conform with BCO-DMO naming conventions
– blank values in this dataset are displayed as “nd” for “no data.” nd is the default missing data identifier in the BCO-DMO system.
– reformatted table so each column is unique
Custom designed pressure chamber used to simulate the high-pressure conditions at the seafloor for this set of experiments.(designed at Rensselaer Polytech Institute, Department of Earth and Environmental Sciences)
No relevant match in BCO-DMO instrument vocabulary.
Used to measure pH.
Used to measure concentrations of ions.
Instruments that quantify carbon, nitrogen and sometimes other elements by combusting the sample at very high temperature and assaying the resulting gaseous oxides. Usually used for samples including organic material.
Experimental conditions applied to experimental units. In comparative experiments, members of the complementary group, the control group, receive either no treatment or a standard treatment.
water pressure at measurement; depth reported as pressure; positive number increasing with water depth
salinity, calculated from the CTD 'primary sensors' of conductivity and temperature, Practical Salinity Scale (PSS-78), dimensionless. Depending on the input source, salinity from the primary sensors can have a variety of names i.e. s0, s00, sal0, sal00.
pH: The measure of the acidity or basicity of an aqueous solution
pH: The measure of the acidity or basicity of an aqueous solution
Concentration of carbonate ion ([CO3]2-) in seawater. Refer to dataset for units of measure.
Calcium (Ca). Concentrations may be reported in parts per million, nanomoles per liter, or other units. Refer to dataset metadata for units.
The saturation state of seawater with respect to aragonite (known as OMEGA_ar) is a measure of the thermodynamic potential for aragonite to form or to dissolve, and is defined as the product of the concentrations of dissolved calcium and carbonate ions in seawater, divided by their product at equilibrium.
| Name | Affiliation | Contact |
|---|---|---|
| Rinat Gabitov | Mississippi State University (MSU) | ✓ |
| Karyn L. Rogers | Mississippi State University (MSU) | ✓ |
| Chiara Borrelli | University of Rochester | |
| Nancy Copley | University of Rochester | |
| Nancy Copley | Woods Hole Oceanographic Institution (WHOI BCO-DMO) |
| Project Title | Biologically induced methane oxidation and precipitation of carbonate minerals: An experimental study |
|---|---|
| Acronym | Deep Sea Geochem CaCO3 |
| URL | https://www.bco-dmo.org/project/743426 |
| Created | August 9, 2018 |
| Modified | September 14, 2018 |
Biologically-mediated CaCO3 precipitation widely occurs in aquatic systems and is often directly linked to the metabolic activity of microorganisms, which could significantly affect the local environment. An example is oxidation of methane and reduction of sulfate mediated by a consortium of Bacteria and Archaea. In order to investigate geochemistry of CaCO3precipitated abiotically and under microbial activity experimental work was initiated. The abiotic experiments were performed at different temperatures and pressures (nitrogen and nitrogen-methane mixture). Further geochemical analyses will allow evaluating of the effect of total pressure and methane partial pressure on the geochemistry of CaCO3. Sulfate reducing bacterial (Desulfovibrio salexigens) was successfully cultured and precipitation experiments on microbially mediated CaCO3 are in progress. This work is relevant to C-DEBI Research Theme I (Activity in the Deep Subseafloor Biosphere: function & rates of global biogeochemical processes) because carbonate growth rate is linked to microbial activity and the rate of methane oxidation.
| Name | Affiliation | Role |
|---|---|---|
| Rinat Gabitov | Mississippi State University (MSU) | Principal Investigator |
| Karyn L. Rogers | Rensselaer Polytechnic Institute (RPI) | Co-Principal Investigator |
| Chiara Borrelli | University of Rochester | Contact |
| Project Title | Biologically induced methane oxidation and precipitation of carbonate minerals: An experimental study |
|---|---|
| Acronym | Deep Sea Geochem CaCO3 |
| URL | https://www.bco-dmo.org/project/743426 |
| Created | August 9, 2018 |
| Modified | September 14, 2018 |
Biologically-mediated CaCO3 precipitation widely occurs in aquatic systems and is often directly linked to the metabolic activity of microorganisms, which could significantly affect the local environment. An example is oxidation of methane and reduction of sulfate mediated by a consortium of Bacteria and Archaea. In order to investigate geochemistry of CaCO3precipitated abiotically and under microbial activity experimental work was initiated. The abiotic experiments were performed at different temperatures and pressures (nitrogen and nitrogen-methane mixture). Further geochemical analyses will allow evaluating of the effect of total pressure and methane partial pressure on the geochemistry of CaCO3. Sulfate reducing bacterial (Desulfovibrio salexigens) was successfully cultured and precipitation experiments on microbially mediated CaCO3 are in progress. This work is relevant to C-DEBI Research Theme I (Activity in the Deep Subseafloor Biosphere: function & rates of global biogeochemical processes) because carbonate growth rate is linked to microbial activity and the rate of methane oxidation.
| Name | Affiliation | Role |
|---|---|---|
| Rinat Gabitov | Mississippi State University (MSU) | Principal Investigator |
| Karyn L. Rogers | Rensselaer Polytechnic Institute (RPI) | Co-Principal Investigator |
| Chiara Borrelli | University of Rochester | Contact |
| URL | https://www.bco-dmo.org/dataset/661659 |
|---|---|
| Download URL | https://www.bco-dmo.org/dataset/661659/data/download |
| Media Type | text/tab-separated-values |
| Created | October 14, 2016 |
| Modified | June 12, 2019 |
| State | Final no updates expected |
| Brief Description | Sulfate reduction energetics at Main Endeavor grotto chimney. |
Tables and Figures referenced in the acquisition description are found in the paper Frank et al., 2015
For each independent treatment, aliquots of 7.5 mL flange slurry (approx. 29 g wet weight and 20 g dry weight) were transferred into Balch tubes in an anaerobic chamber, and supplemented with 15 mL of sterile artificial seawater media designed to mimic the geochemical conditions within a hydrothermal flange (400 mM NaCl, 25 mM KCl, 30 mM CaCl2, 2.3 mM NaHCO3, 14 mM NaSO42-, 1 mM H2S, and 50 uM dissolved organic carbon – consisting of equimolar proportions 10 uM of pyruvate, citrate, formate, acetate, lactate) under a pure nitrogen headspace.
Concentrations of sulfide, sulfate and dissolved organic carbon (DOC) were varied independently to investigate concentration dependent effects on the rates of SR. The range of experimental conditions tested was determined from previously published concentration profiles of aqueous species modeled as functions of temperature and position within the Grotto vent structure (Tivey, 2004). Concentrations were varied by orders of magnitude within the modeled ranges to simulate conditions representative of different mixing regimes between seawater and vent fluid (Table 1). The range of DOC (which we approximate as a mix of pyruvate, citrate, formate, acetate, lactate – most of which have been identified to varying degrees within vent fluid and are known carbon sources for heterotrophic SR in culture) concentrations tested were based on the average DOC concentrations measured within diffuse fluids at the Main Endeavor Field (Lang et al., 2006; Lang et al., 2010). Hydrogen sulfide was present as H2S (pKa in seawater of 6.60) across all the conditions tested (Amend & Shock, 2001). Incubations were carried out at pH 4 (to simulate the pH of end-member Grotto vent fluid and the average calculated pH of mixed fluids in highly reduced zones within the flange; Tivey 2004) as well as pH 6 (representative of the calculated pH in fluid mixing zones; Tivey 2004). All the results are presented and discussed in the context of the initial measured media conditions.
Tables and Figures referenced in the processing description are found in the paper Frank et al., 2015
Potential energy yields of the different metabolisms available in the incubations depend on temperature and fluid compositions. To quantify the energy yield from heterotrophic sulfate reduction (Table 2) in each incubation values of overall Gibbs energy () were calculated according to:
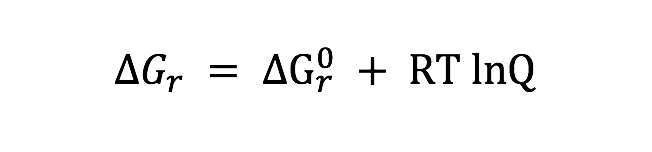
where is the standard Gibbs energy of reaction at in situ temperature and pressure conditions, R is the gas constant, T is the temperature (Kelvin), and Q is the activity product, defined as
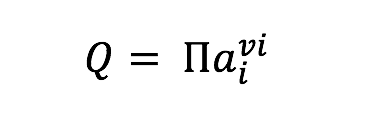
where ai represents the activity of the ith species and vi is the stoichiometric reaction coefficient, which is positive for products and negative for reactants. Values of were calculated at 1 bar and incubation temperatures using the geochemical software package SUPCRT92 (Johnson et al., 1992) and additional thermodynamic data from (Shock, 1995). Activities of aqueous species were calculated using the geochemical speciation program EQ3 (Wolery, 1992) based on the media composition described in section 2.2 and Table 1, with additional data from previously published work (Shock, 1995; Shock & Koretsky, 1993). For concentrations equal to zero, a value of 10-13 mol/kg was used as input. Resulting aqueous activities were used to calculate values of normalized for the number of electrons transferred in the redox for the reactions in Table 2. These reflect the metabolic energy available at the start of each incubation experiment for the complete oxidation of each organic acid, metabolisms that are documented among known sulfate reducers (Amend and Shock, 2001). Furthermore, to calculate the energy density in each incubation (as in Amend et al., 2011), it was assumed that the amended organic acids were the limiting reactant for all experiments when sulfate concentrations were in excess of 1 mM; otherwise sulfate was assumed to be limiting. While some sulfate reducers are known to produce carboxylic acid and alcohol intermediates, incomplete oxidation reactions were not considered here, as the goal of these calculations was to generate a broad understanding of sulfate reduction energetics, and not the metabolic potential for a particular species. Such an approach is common when comparing microbial metabolisms independent of species-specific pathways (e.g. Amend et al., 2004; Rogers & Amend, 2006; Skoog et al., 2007), although it should be noted that incomplete oxidation (fermentation) generally yields much less energy than complete oxidation (Rogers & Amend, 2006; Skoog et al., 2007).
To account for potential interactions between chimney-derived trace metals and amended sulfide, the saturation states of sulfide minerals were calculated as part of the initial fluid speciation. Using reported concentrations of relevant trace metals (Fe, Zn, Cu, etc.) in end-member Grotto hydrothermal fluid (Butterfield et al., 1994), maximum aqueous activities of trace metals were calculated with the EQ3 geochemical speciation program (EQ3/6 1998; EQ3NR 1998). Several sulfide minerals commonly found in hydrothermal chimneys (e.g. pyrite, chalcocite, sphalerite) were supersaturated under incubation conditions, particularly for incubations with high concentrations of amended sulfide. The irreversible abiotic precipitation of mineral sulfides has the potential to draw down aqueous sulfide concentrations and impact sulfate reductions rates. Therefore, the geochemical reaction path program EQ6 (EQ3/6 1998; EQ6 1998) was used to constrain fluid compositions to equilibrium with these minerals phases. Using the single point model in EQ6, the Gibbs energy of the system was allowed to reach local minima by mineral precipitation, however redox reactions among carbon and sulfur species was suppressed with a custom thermodynamic database. The resulting fluid compositions were used to calculate metabolic reaction energetics as well as to evaluate the potential effects of metal speciation on sulfate reduction rates.
BCO-DMO Data Processing Notes:
-reformatted column names to comply with BCO-DMO standards
-filled in all blank cells with nd
-removed spaces and replaced with underscores
pH sensor
DOC was measured
Used aboard ship and in lab
A device on shipboard or in the laboratory that holds water samples under controlled conditions of temperature and possibly illumination.
concentration of sulfide
Concentration of sulfate (SO4) per unit volume
Concentration of bicarbonate ion ([HCO3]-) in seawater. Refer to dataset for units of measure.
K (potassium) concentration. May be reported in parts per million, nanomoles/Liter, or other units. Refer to dataset metadata for units.
Calcium (Ca). Concentrations may be reported in parts per million, nanomoles per liter, or other units. Refer to dataset metadata for units.
Sodium (Na). Concentrations may be reported in parts per million, nanomoles per liter, or other units. Refer to dataset metadata for units.
pH: The measure of the acidity or basicity of an aqueous solution
Temperature in degrees C of a sample or other item. A generic temperature measurement.
Note: This is NOT water temp or sea surface temp
Carbon limiting total energy available in bottle for sulfate reduction
Latitude
latitude, in decimal degrees, North is positive, negative denotes South; Reported in some datasets as degrees, minutes
| Name | Affiliation | Contact |
|---|---|---|
| Peter R. Girguis | Harvard University | |
| Karyn L. Rogers | Harvard University | |
| Kiana L. Frank | Rensselaer Polytechnic Institute (RPI) | |
| Hannah Ake | Rensselaer Polytechnic Institute (RPI) | |
| Shannon Rauch | University of Hawaii at Manoa (SOEST) | ✓ |
| Hannah Ake | Woods Hole Oceanographic Institution (WHOI BCO-DMO) |
| Project Title | Characterizing the distribution and rates of microbial sulfate reduction at Middle Valley hydrothermal vents |
|---|---|
| Acronym | Middle Valley Vents |
| URL | https://www.bco-dmo.org/project/626603 |
| Created | November 17, 2015 |
| Modified | November 19, 2015 |
This project characterizes rates of microbially mediated sulfate reduction from three distinct hydrothermal vents in the Middle Valley vent field along the Juan de Fuca Ridge, as well as assessments of bacterial and archaeal diversity, estimates of total biomass and the abundance of functional genes related to sulfate reduction, and in situ geochemistry. Maximum rates of sulfate reduction occurred at 90°C in all three deposits. Pyrosequencing and functional gene abundance data reveal differences in both biomass and community composition among sites, including differences in the abundance of known sulfate reducing bacteria. The abundance of sequences for Thermodesulfovibro-like organisms and higher sulfate reduction rates at elevated temperatures, suggests that Thermodesulfovibro-like organisms may play a role in sulfate reduction in warmer environments. The rates of sulfate reduction observed suggest that – within anaerobic niches of hydrothermal deposits – heterotrophic sulfate reduction may be quite common and might contribute substantially to secondary productivity, underscoring the potential role of this process in both sulfur and carbon cycling at vents.
This project was funded, in part, by a C-DEBI Graduate Student Fellowship.
| Name | Affiliation | Role |
|---|---|---|
| Peter R. Girguis | Harvard University | Principal Investigator |
| Kiana L. Frank | University of Hawaii at Manoa (SOEST) | Contact |
The ability of microorganisms to withstand long periods with extremely low energy input has gained increasing scientific attention in recent years. Starvation experiments in the laboratory have shown that a phylogenetically wide range of microorganisms evolve fitness-enhancing genetic traits within weeks of incubation under low-energy stress. Studies on natural environments that are cut off from new energy supplies over geologic time scales, such as deeply buried sediments, suggest that similar adaptations might mediate survival under energy limitation in the environment. Yet, the extent to which laboratory-based evidence of starvation survival in pure or mixed cultures can be extrapolated to sustained microbial ecosystems in nature remains unclear. In this review, we discuss past investigations on microbial energy requirements and adaptations to energy limitation, identify gaps in our current knowledge, and outline possible future foci of research on life under extreme energy limitation.
I am requesting funds to visit the laboratory of Dr. Kallmeyer at the University of Potsdam and to work with him and his graduate student, Patrick Sauer, to adapt his high-temperature, high-pressure system to high-temperature, high-pressure deep-sea microbes, and extend the pressure limit to shallow crustal pressure regimes (~100 MPa). The goals of this visit are to (1) test the current high-pressure, high-temperature equipment with a known deep-sea thermophile, Archaeoglobus fulgidus; (2) develop an improved design that increases working pressures to ~100 MPa and includes the addition of fixative to subsamples prior to decompression; and (3) outline future research collaborations between Dr. Kallmeyer, Dr. Rogers and Mr. Sauer. The requested travel funds will solidify a collaboration between two C- DEBI affiliates, including a predoctoral student, and jumpstart a number of future deep biosphere research projects. The transfer of knowledge regarding the high-temperature, high-pressure growth system will greatly expand our ability to investigate the limits of microbial life in the subsurface, and sharing equipment design is one of the key motivations of the C-DEBI community.
Biologically-mediated CaCO3 precipitation widely occurs in aquatic systems and is often directly linked to the metabolic activity of microorganisms, which could significantly affect the local environment. An example is oxidation of methane and reduction of sulfate mediated by a consortium of Bacteria and Archaea. In order to investigate geochemistry of CaCO3 precipitated abiotically and under microbial activity experimental work was initiated. The abiotic experiments were performed at different temperatures and pressures (nitrogen and nitrogen-methane mixture). Further geochemical analyses will allow evaluating of the effect of total pressure and methane partial pressure on the geochemistry of CaCO3. Sulfate reducing bacterial (Desulfovibrio salexigens) was successfully cultured and precipitation experiments on microbially mediated CaCO3 are in progress. This work is relevant to C-DEBI Research Theme I (Activity in the Deep Subseafloor Biosphere: function & rates of global biogeochemical processes) because carbonate growth rate is linked to microbial activity and the rate of methane oxidation.
Seafloor hydrothermal activity dominates heat and chemical exchange between seawater and ocean rocks, however the extent to which diffuse venting contributes to these processes is poorly constrained. The low-temperature, ultra-diffuse venting and extensive iron mats at the FeMO Deep (~0.2°C anomaly) and Shinkai Deep (~6°C anomaly) sites on the flanks of Loihi Seamount provides a unique opportunity to study Fe, C, N, and S cycling in modern analogues to ancient seafloor hematitic formations that are preserved in the rock record. While aerobic microbial activity near the mat surface is responsible for much of the iron deposition, these microaerophiles are limited to the uppermost suboxic zone. Most of the mat thickness is characterized by anoxic conditions, and we hypothesize that anaerobic metabolisms dominate in these zones. Here we propose preliminary studies to characterize this cryptic anaerobic community. We will use isotope-tracer incubation experiments, functional gene analyses and a silver sulfide capture technique to quantify S-, N-, and C-based microbial metabolisms in the anoxic mat interior. Coupled to our ongoing geochemical and bioenergetic efforts, this work will lead to a coupled microbial/geochemical model at FeMO Deep style mats that elucidates the role these systems play in global biogeochemical cycles.