Last Modified: June 12, 2019
accession
NCBI accession number
Database identifier assigned by repository and linked to GenBank or other repository.
| URL | https://www.bco-dmo.org/dataset/717352 |
|---|---|
| Download URL | https://www.bco-dmo.org/dataset/717352/data/download |
| Media Type | text/tab-separated-values |
| Created | October 19, 2017 |
| Modified | June 12, 2019 |
| State | Final no updates expected |
| Brief Description | Lake surface sediment and submarine canyon sediment bacterial V1-V3 iTag sequence libraries. |
Submarine Canyon Sediment Bacteria:
A ~35cm multicore was collected from La Jolla submarine canyon and sectioned using pre-sterilized tools shipboard at 1cm intervals for preservation at -80 deg C. DNA was extracted from ~2g subsamples using the MoBio PowerSoil DNA kit with an additional heating step applied during lysis. Sequence libraries were assembled for select bulk sediment samples for the v1–v3 region of the 16S rRNA gene using Eubacteria-specific primers (Muyzer et al. 1993) on the Illumina MiSeq platform (Bartram et al. 2011) at the University of Minnesota Genomics Center (UMGC).
Lake Surface Sediment Bacteria (excerpt from Harrison et al., 2016):
A 1.5 m hollow steel wedge was filled with a mixture of dry ice pellets and isopropyl alcohol and then lowered into the sediment. The coring device was held in place for 15 min and then raised back to the surface, where the dry ice slurry was poured off and unfrozen mud scraped off the sides. The device was then filled with lake surface water, and the frozen core slabs levered off the sides. The recovered frozen cores were wrapped in aluminum foil and placed on dry ice for transport. In the laboratory, the cores were placed on cardboard and kept frozen by regular contact with dry ice pellets. The sampling plane (facing away from the coring device) was smoothed down using a hand-held electric wood planer and utility knives until undisturbed laminae were clearly resolved. Core sections were selected by visual inspection on the basis of lamina thickness and continuity across the sampling plane.
Laminae were excised with pre-sterilized utility knife blades from the top 9 cm of the Twin Lake freeze core: five corresponding to white layers deposited during spring/summer blooms and four corresponding to winter deposition of organic matter and terrigenous particles. 0.2 g of material was collected from the sampling plane as described above, beginning at the lamina base and removing material no greater than 1 mm above that level.
From the Lake McCarrons freeze core, five spring/summer laminae were collected from sub-sampled portions of the 16–24 cm depth range by sequentially exposing the z-plane surface of the horizon with sterilized utility knife blades.”
Additional 5 mm frozen sediment wedges cross-cutting sediment laminae were taken at 2 cm intervals between 0 cm and 30 cm depth and 5 cm intervals from 30 cm to 65 cm, using pre-sterilized instruments. Subsamples were stored at -80 deg C for later extraction. An additional ambient-temperature piston core was collected from Lake McCarrons sediment at a position roughly adjacent (within 5 m) to the freeze core site and subsampled on shore within 2 h after removal. The presence and vertical migration of gas bubbles was observed in core sediment. 2 cc sediment aliquots were collected at 10 cm depth intervals starting at 2 cm beneath the sediment-water interface through predrilled windows, excluding the exterior 1 cm of material in contact with the coring tube. Subsamples were stored at -80 deg C.
DNA extraction, amplification and sequencing
Depth intervals corresponding to ambient core subsamples and excised laminae were used to select among the high-frequency bulk sediment samples of the McCarrons core. DNA was extracted from all sediment subsamples with a MoBio Soil DNA kit with an additional heating step added to the lysing step of manufacturer protocols (Harrison and Orphan 2012).
Sequence libraries were assembled for select bulk sediment samples for the v1–v3 region of the 16S rRNA gene using Eubacteria-specific primers (Muyzer et al. 1993) on the Illumina MiSeq platform (Bartram et al. 2011).
BCO-DMO Data Processing Notes:
– information was compiled from the submitted metadata forms to form this table of data.
– columns that were created for table: accession, NCBI_link, depth, lat, lon, location, deployment, description
NCBI accession number
NCBI accession link
Link to an external data entry.
Water sample depth
Observation/sample depth below the sea surface. Units often reported as: meters, feet.
When used in a JGOFS/GLOBEC dataset the depth is a best estimate; usually but not always calculated from pressure; calculated either from CTD pressure using Fofonoff and Millard (1982; UNESCO Tech Paper #44) algorithm adjusted for 1980 equation of state for seawater (EOS80) or simply equivalent to nominal depth as recorded during sampling if CTD pressure was unavailable.
Latitude
latitude, in decimal degrees, North is positive, negative denotes South; Reported in some datasets as degrees, minutes
Longitude
longitude, in decimal degrees, East is positive, negative denotes West; Reported in some datsets as degrees, minutes
Deployment name and number
deployment number; often associated with sediment trap deployments
Description of NCBI accession
brief description, open ended, specific to the data set in which it appears
| Name | Affiliation | Contact |
|---|---|---|
| Benjamin Kimball Harrison | Central Michigan University | ✓ |
| Jake V. Bailey | Central Michigan University | ✓ |
| Hannah Ake | University of Minnesota, Twin Cities (UMTC) | |
| Woods Hole Oceanographic Institution (WHOI BCO-DMO) |
| Project Title | Buried alive: Microbial responses to sediment flux with implications for the deep biosphere |
|---|---|
| Acronym | Buried Alive |
| URL | https://www.bco-dmo.org/project/653726 |
| Created | August 10, 2016 |
| Modified | August 10, 2016 |
Previous investigation of the microbial diversity of unconsolidated marine sediments has not yet constrained the importance of cell transport associated with physical processes of sedimentation and fluid advection. Organisms must migrate in order to maintain position with respect to geochemical gradients, and many marine bacteria exhibit chemotactic behavior to optimize their position. However, certain microorganisms in subsurface environments form persistent attachments to solid particles or are non-motile, leading to the differential burial of a subset of active cells (i.e. what is preserved may differ from what is—or was—active at a given horizon). Deep sedimentary horizons may inherit a microbial community that fails to maintain its optimum position with respect to geochemical profiles, and the deep biosphere ultimately is composed of cells that survive this transition.
We seek to describe the permanence and overprinting of molecular signatures of microbial communities across discrete horizons associated with rapid sedimentation (e.g. turbidite emplacement), changes in bottom-water geochemistry, and depositional unconformities. This work involved the collection and study of sediment cores from 3 lakes in the Minneapolis-St. Paul Metropolitan, an intact turbidite profile from La Jolla Canyon on the California Borderland, and select core samples from a marine transgressive sequence drilled by IODP leg 337 off the Shimokita Peninsula, Japan. These core samples provided a key opportunity to study microbial relationships to sedimentology at high resolution. Preliminary work suggests microbial community signatures retain evidence of cell displacement resulting from sediment disturbance, and distinct patterns in diversity are not overprinted on decadal timescales.
This work was supported through a C-DEBI postdoctoral fellowship.
| Name | Affiliation | Role |
|---|---|---|
| Benjamin Kimball Harrison | University of Minnesota, Twin Cities (UMTC) | Lead Principal Investigator |
| Jake V. Bailey | University of Minnesota, Twin Cities (UMTC) | Co-Principal Investigator |
| URL | https://www.bco-dmo.org/dataset/709573 |
|---|---|
| Download URL | https://www.bco-dmo.org/dataset/709573/data/download |
| Media Type | text/tab-separated-values |
| Created | July 24, 2017 |
| Modified | March 26, 2019 |
| State | Final no updates expected |
| Brief Description | Oxygen consumption rates/zero valent iron dissolution of FeOB with kanamycin addition - replacement samples |
These data were collected by placing each strain in a 100 mL serum vial with 6 mL of their standard, published media with 30 mg zero valent iron as a source of Fe(II). The headspace was filled with a gas mix of 8% oxygen/10% carbon dioxide/82% nitrogen by using bottled gas mixes and a regulator to flush the headspace without over pressurization. Prior to sealing the serum vials, a Presens OPTODE dot (sensor) was placed inside the vial, allowing non-invasive gas sampling of the changes in O2 in the headspace. A Presens four channel system was used to measure changes in oxygen concentration in real-time in each bottle. A total of four channels were measured during each experiment: channels 1 through 3 are the biological treatments and channel 4 was a kill control (microbes were by placing on a heat block at 100 degrees C for 5 minutes). After 3 days of incubation, the concentration of Fe(II) in the media was measured by ferrozine assay. Old media was then removed and replaced with fresh media containing 30 ng/ml kanamycin to prevent growth of remaining cells. The headspace was again filled with the same gas mix and oxygen concentrations were measured in real-time in each vial. Fe(II) concentration was determined daily by ferrozine assay for 3 more days.
Data was acquired using PreSens Measurement Studio 2 RC4 v0.5.6039.20506
Oxygen concentration readings cut out for PV-1 after approximately 4 days – will be fixed in future experiment.
BCO-DMO Data Processing Notes:
-Reformatted column names to comply with BCO-DMO standards
-Reformatted dates from mm/dd/yy to yyyy/mm/dd
-Replaced spaces with underscores
-Replaced N/A with nd
Used with air saturated water and 100% nitrogen gas.
unique sample identification or number; any combination of alpha numeric characters; precise definition is file dependent
date; generally reported in GMT as YYYYMMDD (year; month; day); also as MMDD (month; day); EqPac dates are local Hawaii time. ISO_Date format is YYYY-MM-DD (http://www.iso.org/iso/home/standards/iso8601.htm)
Experimental conditions applied to experimental units. In comparative experiments, members of the complementary group, the control group, receive either no treatment or a standard treatment.
Oxygen; dissolved; reported in units of micromoles/liter
Date and time (UTC) formatted to ISO8601 standard. T indicates start of time string; Z indicates UTC.
Date/Time (UTC) ISO formatted
This standard is based on ISO 8601:2004(E) and takes on any of the following forms:
2009-08-30T09:05:00[.xx] (local time)
2009-08-30T14:05:00[.xx]Z (UTC time)
2009-08-30T14:05:00[.xx]-05:00
The dashes and the colons can be dropped.
The T can also be dropped "by mutual agreement", but one needs the trailing Z if the time is UTC.
Sample ISO_datetime_utc:
2009-08-30T14:05:00[.xx]Z (UTC time)
| Name | Affiliation | Contact |
|---|---|---|
| Peter R. Girguis | Harvard University | |
| David Emerson | Harvard University | |
| Jacob Cohen | Bigelow Laboratory for Ocean Sciences | |
| Hannah Ake | Bigelow Laboratory for Ocean Sciences | |
| Hannah Ake | Woods Hole Oceanographic Institution (WHOI BCO-DMO) |
| Project Title | Collaborative Research: The Role of Iron-oxidizing Bacteria in the Sedimentary Iron Cycle: Ecological, Physiological and Biogeochemical Implications |
|---|---|
| Acronym | SedimentaryIronCycle |
| URL | https://www.bco-dmo.org/project/544584 |
| Created | January 8, 2015 |
| Modified | June 1, 2018 |
Iron is a critical element for life that serves as an essential trace element for eukaryotic organisms. It is also able to support the growth of a cohort of microbes that can either gain energy for growth via oxidation of ferrous (Fe(II)) to ferric (Fe(III)) iron, or by utilizing Fe(III) for anaerobic respiration coupled to oxidation of simple organic matter or H2. This coupled process is referred to as the microbial iron cycle. One of the primary sources of iron to the ocean comes from dissolved iron (dFe) that is produced through oxidation and reduction processes in the sediment where iron is abundant. The dFe is transported into the overlaying water where it is an essential nutrient for phytoplankton responsible for primary production in the world’s oceans. In fact, iron limitation significantly impacts production in as much as a third of the world’s open oceans. The basic geochemistry of this process is understood; however important gaps exist in our knowledge about the details of how the iron cycle works, and how critical a role bacteria play in it.
Intellectual Merit. Conventional wisdom holds that most of the iron oxidation in sediments is abiological, as a result of the rapid kinetics of chemical iron oxidation in the presence of oxygen. This proposal aims to question this conventional view and enhance our understanding of the microbes involved in the sedimentary iron cycle, with an emphasis on the bacteria that catalyze the oxidation of iron. These Fe-oxidizing bacteria (FeOB) utilize iron as a sole energy source for growth, and are autotrophic. They were only discovered in the ocean about forty-five years ago, and are now known to be abundant at hydrothermal vents that emanate ferrous-rich fluids. More recently, the first evidence was published that they could inhabit coastal sediments, albeit at reduced numbers, and even be abundant in some continental shelf sediments. These habitats are far removed from hydrothermal vents, and reveal the sediments may be an important habitat for FeOB that live on ferrous iron generated in the sediment. This begs the question: are FeOB playing an important role in the oxidative part of the sedimentary Fe-cycle? One important attribute of FeOB is their ability to grow at very low levels of O2, an essential strategy for them to outcompete chemical iron oxidation. How low a level of O2 can sustain them, and how this might affect their distribution in sediments is unknown. In part, this is due to the technical challenges of measuring O2 concentrations and dynamics at very low levels; yet these concentrations could be where FeOB flourish. The central hypothesis of this proposal is that FeOB are more common in marine sedimentary environments than previously recognized, and play a substantive role in governing the iron flux from the sediments into the water column by constraining the release of dFe from sediments. A set of experimental objectives are proposed to test this. A survey of near shore regions in the Gulf of Maine, and a transect along the Monterey Canyon off the coast of California will obtain cores of sedimentary muds and look at the vertical distribution of FeOB and putative Fe-reducing bacteria using sensitive techniques to detect their presence and relative abundance. Some of these same sediments will be used in a novel reactor system that will allow for precise control of O2 levels and iron concentration to measure the dynamics of the iron cycle under different oxygen regimens. Finally pure cultures of FeOB with different O2 affinities will be tested in a bioreactor coupled to a highly sensitive mass spectrometer to determine the lower limits of O2 utilization for different FeOB growing on iron, thus providing mechanistic insight into their activity and distribution in low oxygen environments.
Broader Impacts. An important impact of climate change on marine environments is a predicted increase in low O2 or hypoxic zones in the ocean. Hypoxia in association with marine sediments will have a profound influence on the sedimentary iron cycle, and is likely to lead to greater inputs of dFe into the ocean. In the longer term, this increase in dFe flux could alleviate iron-limitation in some regions of the ocean, thereby enhancing the rate of CO2-fixation and draw down of CO2 from the atmosphere. This is one important reason for developing a better understanding of microbial control of sedimentary iron cycle. This project will also provide training to a postdoctoral scientist, graduate students and undergraduates. This project will contribute to a student initiated exhibit, entitled ‘Iron and the evolution of life on Earth’ at the Harvard Museum of Natural History providing a unique opportunity for undergraduate training and outreach.
| Name | Affiliation | Role |
|---|---|---|
| David Emerson | Bigelow Laboratory for Ocean Sciences | Principal Investigator |
| Peter R. Girguis | Harvard University | Principal Investigator |
| David Johnson | Harvard University | Co-Principal Investigator |
| URL | https://www.bco-dmo.org/dataset/709440 |
|---|---|
| Download URL | https://www.bco-dmo.org/dataset/709440/data/download |
| Media Type | text/tab-separated-values |
| Created | July 21, 2017 |
| Modified | March 26, 2019 |
| State | Final no updates expected |
| Brief Description | Fe(II) Concentrations over time in iron oxidizing bacteria cultures. |
These data were collected by placing each strain in a 100 mL serum vial with 6 mL of their standard, published media with 30 mg zero valent iron as a source of Fe(II). The headspace was filled with a gas mix of 8% oxygen/10% carbon dioxide/82% nitrogen by using bottled gas mixes and a regulator to flush the headspace without overpressurization. Each vial was sampled once per day and the Fe(II) concentration was determined by ferrozine assay.
BCO-DMO Data Processing Notes:
-Reformatted column names to comply with BCO-DMO standards
-Reformatted dates from mm/dd/yy to yyyy/mm/dd
-Replaced spaces with underscores
Used to measure ferrozine assay
An instrument used to measure the relative absorption of electromagnetic radiation of different wavelengths in the near infra-red, visible and ultraviolet wavebands by samples.
unique sample identification or number; any combination of alpha numeric characters; precise definition is file dependent
date; generally reported in GMT as YYYYMMDD (year; month; day); also as MMDD (month; day); EqPac dates are local Hawaii time. ISO_Date format is YYYY-MM-DD (http://www.iso.org/iso/home/standards/iso8601.htm)
Iron (Fe). Concentrations may be reported in parts per million, nanomoles per liter, or other units. Refer to dataset metadata for units.
| Name | Affiliation | Contact |
|---|---|---|
| Peter R. Girguis | Harvard University | |
| David Emerson | Harvard University | |
| Jacob Cohen | Bigelow Laboratory for Ocean Sciences | |
| Hannah Ake | Bigelow Laboratory for Ocean Sciences | |
| Hannah Ake | Woods Hole Oceanographic Institution (WHOI BCO-DMO) |
| Project Title | Collaborative Research: The Role of Iron-oxidizing Bacteria in the Sedimentary Iron Cycle: Ecological, Physiological and Biogeochemical Implications |
|---|---|
| Acronym | SedimentaryIronCycle |
| URL | https://www.bco-dmo.org/project/544584 |
| Created | January 8, 2015 |
| Modified | June 1, 2018 |
Iron is a critical element for life that serves as an essential trace element for eukaryotic organisms. It is also able to support the growth of a cohort of microbes that can either gain energy for growth via oxidation of ferrous (Fe(II)) to ferric (Fe(III)) iron, or by utilizing Fe(III) for anaerobic respiration coupled to oxidation of simple organic matter or H2. This coupled process is referred to as the microbial iron cycle. One of the primary sources of iron to the ocean comes from dissolved iron (dFe) that is produced through oxidation and reduction processes in the sediment where iron is abundant. The dFe is transported into the overlaying water where it is an essential nutrient for phytoplankton responsible for primary production in the world’s oceans. In fact, iron limitation significantly impacts production in as much as a third of the world’s open oceans. The basic geochemistry of this process is understood; however important gaps exist in our knowledge about the details of how the iron cycle works, and how critical a role bacteria play in it.
Intellectual Merit. Conventional wisdom holds that most of the iron oxidation in sediments is abiological, as a result of the rapid kinetics of chemical iron oxidation in the presence of oxygen. This proposal aims to question this conventional view and enhance our understanding of the microbes involved in the sedimentary iron cycle, with an emphasis on the bacteria that catalyze the oxidation of iron. These Fe-oxidizing bacteria (FeOB) utilize iron as a sole energy source for growth, and are autotrophic. They were only discovered in the ocean about forty-five years ago, and are now known to be abundant at hydrothermal vents that emanate ferrous-rich fluids. More recently, the first evidence was published that they could inhabit coastal sediments, albeit at reduced numbers, and even be abundant in some continental shelf sediments. These habitats are far removed from hydrothermal vents, and reveal the sediments may be an important habitat for FeOB that live on ferrous iron generated in the sediment. This begs the question: are FeOB playing an important role in the oxidative part of the sedimentary Fe-cycle? One important attribute of FeOB is their ability to grow at very low levels of O2, an essential strategy for them to outcompete chemical iron oxidation. How low a level of O2 can sustain them, and how this might affect their distribution in sediments is unknown. In part, this is due to the technical challenges of measuring O2 concentrations and dynamics at very low levels; yet these concentrations could be where FeOB flourish. The central hypothesis of this proposal is that FeOB are more common in marine sedimentary environments than previously recognized, and play a substantive role in governing the iron flux from the sediments into the water column by constraining the release of dFe from sediments. A set of experimental objectives are proposed to test this. A survey of near shore regions in the Gulf of Maine, and a transect along the Monterey Canyon off the coast of California will obtain cores of sedimentary muds and look at the vertical distribution of FeOB and putative Fe-reducing bacteria using sensitive techniques to detect their presence and relative abundance. Some of these same sediments will be used in a novel reactor system that will allow for precise control of O2 levels and iron concentration to measure the dynamics of the iron cycle under different oxygen regimens. Finally pure cultures of FeOB with different O2 affinities will be tested in a bioreactor coupled to a highly sensitive mass spectrometer to determine the lower limits of O2 utilization for different FeOB growing on iron, thus providing mechanistic insight into their activity and distribution in low oxygen environments.
Broader Impacts. An important impact of climate change on marine environments is a predicted increase in low O2 or hypoxic zones in the ocean. Hypoxia in association with marine sediments will have a profound influence on the sedimentary iron cycle, and is likely to lead to greater inputs of dFe into the ocean. In the longer term, this increase in dFe flux could alleviate iron-limitation in some regions of the ocean, thereby enhancing the rate of CO2-fixation and draw down of CO2 from the atmosphere. This is one important reason for developing a better understanding of microbial control of sedimentary iron cycle. This project will also provide training to a postdoctoral scientist, graduate students and undergraduates. This project will contribute to a student initiated exhibit, entitled ‘Iron and the evolution of life on Earth’ at the Harvard Museum of Natural History providing a unique opportunity for undergraduate training and outreach.
| Name | Affiliation | Role |
|---|---|---|
| David Emerson | Bigelow Laboratory for Ocean Sciences | Principal Investigator |
| Peter R. Girguis | Harvard University | Principal Investigator |
| David Johnson | Harvard University | Co-Principal Investigator |
| URL | https://www.bco-dmo.org/dataset/709543 |
|---|---|
| Download URL | https://www.bco-dmo.org/dataset/709543/data/download |
| Media Type | text/tab-separated-values |
| Created | July 24, 2017 |
| Modified | March 26, 2019 |
| State | Final no updates expected |
| Brief Description | Oxygen consumption rates/zero valent iron dissolution of FeOB with kanamycin addition - Ferrozine assay |
These data were collected by placing each strain in a 100 mL serum vial with 6 mL of their standard, published media with 30 mg zero valent iron as a source of Fe(II). The headspace was filled with a gas mix of 8% oxygen/10% carbon dioxide/82% nitrogen by using bottled gas mixes and a regulator to flush the headspace without over pressurization. Prior to sealing the serum vials, a Presens OPTODE dot (sensor) was placed inside the vial, allowing non-invasive gas sampling of the changes in O2 in the headspace. A Presens four channel system was used to measure changes in oxygen concentration in real-time in each bottle. A total of four channels were measured during each experiment: channels 1 through 3 are the biological treatments and channel 4 was a kill control (microbes were by placing on a heat block at 100 degrees C for 5 minutes). After 3 days of incubation, the concentration of Fe(II) in the media was measured by ferrozine assay. Old media was then removed and replaced with fresh media containing 30 ng/ml kanamycin to prevent growth of remaining cells. The headspace was again filled with the same gas mix and oxygen concentrations were measured in real-time in each vial. Fe(II) concentration was determined daily by ferrozine assay for 3 more days.
Data was acquired using PreSens Measurement Studio 2 RC4 v0.5.6039.20506
Oxygen concentration readings cut out for PV-1 after approximately 4 days – will be fixed in future experiment.
BCO-DMO Data Processing Notes:
-Reformatted column names to comply with BCO-DMO standards
-Reformatted dates from mm/dd/yy to yyyy/mm/dd
-Replaced spaces with underscores
-Replaced N/A with nd
Used to measure ferrozine assay
An instrument used to measure the relative absorption of electromagnetic radiation of different wavelengths in the near infra-red, visible and ultraviolet wavebands by samples.
Used with air saturated water and 100% nitrogen gas.
unique sample identification or number; any combination of alpha numeric characters; precise definition is file dependent
date; generally reported in GMT as YYYYMMDD (year; month; day); also as MMDD (month; day); EqPac dates are local Hawaii time. ISO_Date format is YYYY-MM-DD (http://www.iso.org/iso/home/standards/iso8601.htm)
Experimental conditions applied to experimental units. In comparative experiments, members of the complementary group, the control group, receive either no treatment or a standard treatment.
Iron (Fe). Concentrations may be reported in parts per million, nanomoles per liter, or other units. Refer to dataset metadata for units.
| Name | Affiliation | Contact |
|---|---|---|
| Peter R. Girguis | Harvard University | |
| David Emerson | Harvard University | |
| Jacob Cohen | Bigelow Laboratory for Ocean Sciences | |
| Hannah Ake | Bigelow Laboratory for Ocean Sciences | |
| Hannah Ake | Woods Hole Oceanographic Institution (WHOI BCO-DMO) |
| Project Title | Collaborative Research: The Role of Iron-oxidizing Bacteria in the Sedimentary Iron Cycle: Ecological, Physiological and Biogeochemical Implications |
|---|---|
| Acronym | SedimentaryIronCycle |
| URL | https://www.bco-dmo.org/project/544584 |
| Created | January 8, 2015 |
| Modified | June 1, 2018 |
Iron is a critical element for life that serves as an essential trace element for eukaryotic organisms. It is also able to support the growth of a cohort of microbes that can either gain energy for growth via oxidation of ferrous (Fe(II)) to ferric (Fe(III)) iron, or by utilizing Fe(III) for anaerobic respiration coupled to oxidation of simple organic matter or H2. This coupled process is referred to as the microbial iron cycle. One of the primary sources of iron to the ocean comes from dissolved iron (dFe) that is produced through oxidation and reduction processes in the sediment where iron is abundant. The dFe is transported into the overlaying water where it is an essential nutrient for phytoplankton responsible for primary production in the world’s oceans. In fact, iron limitation significantly impacts production in as much as a third of the world’s open oceans. The basic geochemistry of this process is understood; however important gaps exist in our knowledge about the details of how the iron cycle works, and how critical a role bacteria play in it.
Intellectual Merit. Conventional wisdom holds that most of the iron oxidation in sediments is abiological, as a result of the rapid kinetics of chemical iron oxidation in the presence of oxygen. This proposal aims to question this conventional view and enhance our understanding of the microbes involved in the sedimentary iron cycle, with an emphasis on the bacteria that catalyze the oxidation of iron. These Fe-oxidizing bacteria (FeOB) utilize iron as a sole energy source for growth, and are autotrophic. They were only discovered in the ocean about forty-five years ago, and are now known to be abundant at hydrothermal vents that emanate ferrous-rich fluids. More recently, the first evidence was published that they could inhabit coastal sediments, albeit at reduced numbers, and even be abundant in some continental shelf sediments. These habitats are far removed from hydrothermal vents, and reveal the sediments may be an important habitat for FeOB that live on ferrous iron generated in the sediment. This begs the question: are FeOB playing an important role in the oxidative part of the sedimentary Fe-cycle? One important attribute of FeOB is their ability to grow at very low levels of O2, an essential strategy for them to outcompete chemical iron oxidation. How low a level of O2 can sustain them, and how this might affect their distribution in sediments is unknown. In part, this is due to the technical challenges of measuring O2 concentrations and dynamics at very low levels; yet these concentrations could be where FeOB flourish. The central hypothesis of this proposal is that FeOB are more common in marine sedimentary environments than previously recognized, and play a substantive role in governing the iron flux from the sediments into the water column by constraining the release of dFe from sediments. A set of experimental objectives are proposed to test this. A survey of near shore regions in the Gulf of Maine, and a transect along the Monterey Canyon off the coast of California will obtain cores of sedimentary muds and look at the vertical distribution of FeOB and putative Fe-reducing bacteria using sensitive techniques to detect their presence and relative abundance. Some of these same sediments will be used in a novel reactor system that will allow for precise control of O2 levels and iron concentration to measure the dynamics of the iron cycle under different oxygen regimens. Finally pure cultures of FeOB with different O2 affinities will be tested in a bioreactor coupled to a highly sensitive mass spectrometer to determine the lower limits of O2 utilization for different FeOB growing on iron, thus providing mechanistic insight into their activity and distribution in low oxygen environments.
Broader Impacts. An important impact of climate change on marine environments is a predicted increase in low O2 or hypoxic zones in the ocean. Hypoxia in association with marine sediments will have a profound influence on the sedimentary iron cycle, and is likely to lead to greater inputs of dFe into the ocean. In the longer term, this increase in dFe flux could alleviate iron-limitation in some regions of the ocean, thereby enhancing the rate of CO2-fixation and draw down of CO2 from the atmosphere. This is one important reason for developing a better understanding of microbial control of sedimentary iron cycle. This project will also provide training to a postdoctoral scientist, graduate students and undergraduates. This project will contribute to a student initiated exhibit, entitled ‘Iron and the evolution of life on Earth’ at the Harvard Museum of Natural History providing a unique opportunity for undergraduate training and outreach.
| Name | Affiliation | Role |
|---|---|---|
| David Emerson | Bigelow Laboratory for Ocean Sciences | Principal Investigator |
| Peter R. Girguis | Harvard University | Principal Investigator |
| David Johnson | Harvard University | Co-Principal Investigator |
| URL | https://www.bco-dmo.org/dataset/661659 |
|---|---|
| Download URL | https://www.bco-dmo.org/dataset/661659/data/download |
| Media Type | text/tab-separated-values |
| Created | October 14, 2016 |
| Modified | June 12, 2019 |
| State | Final no updates expected |
| Brief Description | Sulfate reduction energetics at Main Endeavor grotto chimney. |
Tables and Figures referenced in the acquisition description are found in the paper Frank et al., 2015
For each independent treatment, aliquots of 7.5 mL flange slurry (approx. 29 g wet weight and 20 g dry weight) were transferred into Balch tubes in an anaerobic chamber, and supplemented with 15 mL of sterile artificial seawater media designed to mimic the geochemical conditions within a hydrothermal flange (400 mM NaCl, 25 mM KCl, 30 mM CaCl2, 2.3 mM NaHCO3, 14 mM NaSO42-, 1 mM H2S, and 50 uM dissolved organic carbon – consisting of equimolar proportions 10 uM of pyruvate, citrate, formate, acetate, lactate) under a pure nitrogen headspace.
Concentrations of sulfide, sulfate and dissolved organic carbon (DOC) were varied independently to investigate concentration dependent effects on the rates of SR. The range of experimental conditions tested was determined from previously published concentration profiles of aqueous species modeled as functions of temperature and position within the Grotto vent structure (Tivey, 2004). Concentrations were varied by orders of magnitude within the modeled ranges to simulate conditions representative of different mixing regimes between seawater and vent fluid (Table 1). The range of DOC (which we approximate as a mix of pyruvate, citrate, formate, acetate, lactate – most of which have been identified to varying degrees within vent fluid and are known carbon sources for heterotrophic SR in culture) concentrations tested were based on the average DOC concentrations measured within diffuse fluids at the Main Endeavor Field (Lang et al., 2006; Lang et al., 2010). Hydrogen sulfide was present as H2S (pKa in seawater of 6.60) across all the conditions tested (Amend & Shock, 2001). Incubations were carried out at pH 4 (to simulate the pH of end-member Grotto vent fluid and the average calculated pH of mixed fluids in highly reduced zones within the flange; Tivey 2004) as well as pH 6 (representative of the calculated pH in fluid mixing zones; Tivey 2004). All the results are presented and discussed in the context of the initial measured media conditions.
Tables and Figures referenced in the processing description are found in the paper Frank et al., 2015
Potential energy yields of the different metabolisms available in the incubations depend on temperature and fluid compositions. To quantify the energy yield from heterotrophic sulfate reduction (Table 2) in each incubation values of overall Gibbs energy () were calculated according to:
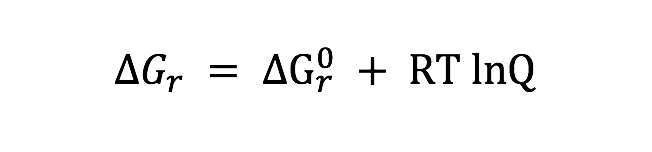
where is the standard Gibbs energy of reaction at in situ temperature and pressure conditions, R is the gas constant, T is the temperature (Kelvin), and Q is the activity product, defined as
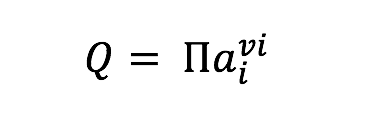
where ai represents the activity of the ith species and vi is the stoichiometric reaction coefficient, which is positive for products and negative for reactants. Values of were calculated at 1 bar and incubation temperatures using the geochemical software package SUPCRT92 (Johnson et al., 1992) and additional thermodynamic data from (Shock, 1995). Activities of aqueous species were calculated using the geochemical speciation program EQ3 (Wolery, 1992) based on the media composition described in section 2.2 and Table 1, with additional data from previously published work (Shock, 1995; Shock & Koretsky, 1993). For concentrations equal to zero, a value of 10-13 mol/kg was used as input. Resulting aqueous activities were used to calculate values of normalized for the number of electrons transferred in the redox for the reactions in Table 2. These reflect the metabolic energy available at the start of each incubation experiment for the complete oxidation of each organic acid, metabolisms that are documented among known sulfate reducers (Amend and Shock, 2001). Furthermore, to calculate the energy density in each incubation (as in Amend et al., 2011), it was assumed that the amended organic acids were the limiting reactant for all experiments when sulfate concentrations were in excess of 1 mM; otherwise sulfate was assumed to be limiting. While some sulfate reducers are known to produce carboxylic acid and alcohol intermediates, incomplete oxidation reactions were not considered here, as the goal of these calculations was to generate a broad understanding of sulfate reduction energetics, and not the metabolic potential for a particular species. Such an approach is common when comparing microbial metabolisms independent of species-specific pathways (e.g. Amend et al., 2004; Rogers & Amend, 2006; Skoog et al., 2007), although it should be noted that incomplete oxidation (fermentation) generally yields much less energy than complete oxidation (Rogers & Amend, 2006; Skoog et al., 2007).
To account for potential interactions between chimney-derived trace metals and amended sulfide, the saturation states of sulfide minerals were calculated as part of the initial fluid speciation. Using reported concentrations of relevant trace metals (Fe, Zn, Cu, etc.) in end-member Grotto hydrothermal fluid (Butterfield et al., 1994), maximum aqueous activities of trace metals were calculated with the EQ3 geochemical speciation program (EQ3/6 1998; EQ3NR 1998). Several sulfide minerals commonly found in hydrothermal chimneys (e.g. pyrite, chalcocite, sphalerite) were supersaturated under incubation conditions, particularly for incubations with high concentrations of amended sulfide. The irreversible abiotic precipitation of mineral sulfides has the potential to draw down aqueous sulfide concentrations and impact sulfate reductions rates. Therefore, the geochemical reaction path program EQ6 (EQ3/6 1998; EQ6 1998) was used to constrain fluid compositions to equilibrium with these minerals phases. Using the single point model in EQ6, the Gibbs energy of the system was allowed to reach local minima by mineral precipitation, however redox reactions among carbon and sulfur species was suppressed with a custom thermodynamic database. The resulting fluid compositions were used to calculate metabolic reaction energetics as well as to evaluate the potential effects of metal speciation on sulfate reduction rates.
BCO-DMO Data Processing Notes:
-reformatted column names to comply with BCO-DMO standards
-filled in all blank cells with nd
-removed spaces and replaced with underscores
pH sensor
DOC was measured
Used aboard ship and in lab
A device on shipboard or in the laboratory that holds water samples under controlled conditions of temperature and possibly illumination.
concentration of sulfide
Concentration of sulfate (SO4) per unit volume
Concentration of bicarbonate ion ([HCO3]-) in seawater. Refer to dataset for units of measure.
K (potassium) concentration. May be reported in parts per million, nanomoles/Liter, or other units. Refer to dataset metadata for units.
Calcium (Ca). Concentrations may be reported in parts per million, nanomoles per liter, or other units. Refer to dataset metadata for units.
Sodium (Na). Concentrations may be reported in parts per million, nanomoles per liter, or other units. Refer to dataset metadata for units.
pH: The measure of the acidity or basicity of an aqueous solution
Temperature in degrees C of a sample or other item. A generic temperature measurement.
Note: This is NOT water temp or sea surface temp
Carbon limiting total energy available in bottle for sulfate reduction
Latitude
latitude, in decimal degrees, North is positive, negative denotes South; Reported in some datasets as degrees, minutes
| Name | Affiliation | Contact |
|---|---|---|
| Peter R. Girguis | Harvard University | |
| Karyn L. Rogers | Harvard University | |
| Kiana L. Frank | Rensselaer Polytechnic Institute (RPI) | |
| Hannah Ake | Rensselaer Polytechnic Institute (RPI) | |
| Shannon Rauch | University of Hawaii at Manoa (SOEST) | ✓ |
| Hannah Ake | Woods Hole Oceanographic Institution (WHOI BCO-DMO) |
| Project Title | Characterizing the distribution and rates of microbial sulfate reduction at Middle Valley hydrothermal vents |
|---|---|
| Acronym | Middle Valley Vents |
| URL | https://www.bco-dmo.org/project/626603 |
| Created | November 17, 2015 |
| Modified | November 19, 2015 |
This project characterizes rates of microbially mediated sulfate reduction from three distinct hydrothermal vents in the Middle Valley vent field along the Juan de Fuca Ridge, as well as assessments of bacterial and archaeal diversity, estimates of total biomass and the abundance of functional genes related to sulfate reduction, and in situ geochemistry. Maximum rates of sulfate reduction occurred at 90°C in all three deposits. Pyrosequencing and functional gene abundance data reveal differences in both biomass and community composition among sites, including differences in the abundance of known sulfate reducing bacteria. The abundance of sequences for Thermodesulfovibro-like organisms and higher sulfate reduction rates at elevated temperatures, suggests that Thermodesulfovibro-like organisms may play a role in sulfate reduction in warmer environments. The rates of sulfate reduction observed suggest that – within anaerobic niches of hydrothermal deposits – heterotrophic sulfate reduction may be quite common and might contribute substantially to secondary productivity, underscoring the potential role of this process in both sulfur and carbon cycling at vents.
This project was funded, in part, by a C-DEBI Graduate Student Fellowship.
| Name | Affiliation | Role |
|---|---|---|
| Peter R. Girguis | Harvard University | Principal Investigator |
| Kiana L. Frank | University of Hawaii at Manoa (SOEST) | Contact |
| URL | https://www.bco-dmo.org/dataset/661557 |
|---|---|
| Download URL | https://www.bco-dmo.org/dataset/661557/data/download |
| Media Type | text/tab-separated-values |
| Created | October 13, 2016 |
| Modified | April 22, 2019 |
| State | Final no updates expected |
| Brief Description | Sulfate reduction rates at Main Endeavor grotto chimney |
Tables and Figures referenced in the acquisition description are found in the paper Frank et al., 2015
Once on board ship, tubeworms and other macrofauna were removed from the samples and the large pieces were broken into more manageable fragments (~10-20 cm3) with a flame-sterilized chisel and sledgehammer, with the user wearing sterile nitrile gloves. Samples were quickly transferred to 0.2 um-filtered anaerobic (nitrogen-sparged) seawater. Samples were further broken down into smaller sizes while in anaerobic water, and subsamples from the interior of the fragments were immediately transferred to gastight jars (Freund Container Inc.) filled with sterile anaerobic seawater containing 2 mM sodium sulfide at pH 6, and stored at 4 degrees celsius for incubations and analyses. The sterile sulfidic seawater in the gastight jars were refreshed periodically during storage at 4 degrees celsius. The majority of the rate experiments (80%) were set up immediately on the ship using freshly collected samples. In parallel, subsamples (~1 cm3) from each flange were preserved aboard ship in glutaraldehyde (2.5% in phosphate buffered saline, PBS, pH 7.0), then prepared for electron microscopy via ethanol dehydration and critical point drying before being sputtered with a thin layer of gold-palladium to improve image resolution. Samples were imaged with a Zeiss model EVO Scanning Electron Microscope (SEM).
Prior to incubation, each flange subsample was pulverized by hand for about one hour to minimize fine-scale geological and microbial heterogeneity and facilitate more accurate experimental replication (akin to slurry experiments in sediments; Fossing & Jørgensen 1989; Weber & Jørgensen 2002; Jørgensen et al. 1992). Specifically, each subsample was pulverized with a flame-sterilized sledgehammer in sterile seawater actively bubbled with nitrogen within an anaerobic chamber. For each independent treatment, aliquots of 7.5 mL flange slurry (approx. 29 g wet weight and 20 g dry weight) were transferred into Balch tubes in an anaerobic chamber, and supplemented with 15 mL of sterile artificial seawater media designed to mimic the geochemical conditions within a hydrothermal flange (400 mM NaCl, 25 mM KCl, 30 mM CaCl2, 2.3 mM NaHCO3, 14 mM NaSO42-, 1 mM H2S, and 50 uM dissolved organic carbon – consisting of equimolar proportions 10 uM of pyruvate, citrate, formate, acetate, lactate) under a pure nitrogen headspace.
Concentrations of sulfide, sulfate and dissolved organic carbon (DOC) were varied independently to investigate concentration dependent effects on the rates of SR. The range of experimental conditions tested was determined from previously published concentration profiles of aqueous species modeled as functions of temperature and position within the Grotto vent structure (Tivey, 2004). Concentrations were varied by orders of magnitude within the modeled ranges to simulate conditions representative of different mixing regimes between seawater and vent fluid (Table 1). The range of DOC (which we approximate as a mix of pyruvate, citrate, formate, acetate, lactate – most of which have been identified to varying degrees within vent fluid and are known carbon sources for heterotrophic SR in culture) concentrations tested were based on the average DOC concentrations measured within diffuse fluids at the Main Endeavor Field (Lang et al., 2006; Lang et al., 2010). Hydrogen sulfide was present as H2S (pKa in seawater of 6.60) across all the conditions tested (Amend & Shock, 2001). Incubations were carried out at pH 4 (to simulate the pH of end-member Grotto vent fluid and the average calculated pH of mixed fluids in highly reduced zones within the flange; Tivey 2004) as well as pH 6 (representative of the calculated pH in fluid mixing zones; Tivey 2004). All the results are presented and discussed in the context of the initial measured media conditions.
Sufficient 35SO42- was added to achieve 15 uCi of activity. Samples were incubated anaerobically for 1, 3 or 7 days at ambient seawater (4 degrees celsius), thermophilic (50 degrees celsius) and hyperthermophilic (90 degrees celsius) temperatures. The range of temperatures considered was representative of different thermal regimes associated with the surface, outer layer and middle regions of hydrothermal chimneys (Tivey 2004; Kormas et al. 2006; Schrenk et al. 2003). Negative controls consisted of samples amended with 28 mM molybdate to inhibit SR (Newport & Nedwell, 1988; Saleh et al., 1964). Three biological replicates were run for each treatment, and two biological replicates for each control.
Upon completion, reactions were quenched with the injection of 5 mL 25% zinc acetate, at pH 8 (i.e. 20-fold excess Zn), and all samples were frozen at -20 degrees celsius for further analysis. 80% of incubations were performed shipboard with freshly collected samples and the remaining 20% of incubations were completed within one year of collection.
To determine SR rates, samples were thawed and the supernatant was removed and filtered through a 0.2 um syringe filter. The homogenized flange that remained in the tube was washed three times with deionized water to remove any remaining sulfate. One gram (wet weight) of flange material was added to 10 mL of a 1:1 ethanol to water solution in the chromium distillation apparatus, and then degassed with nitrogen for 15 minutes to drive the environment anoxic. Hydrogen sulfide gas was evolved after the anaerobic addition of 8 mL of 12 N HCl and 10 mL of 1 M reduced chromium chloride, followed by 3 hours of heating. The resulting hydrogen sulfide gas was carried via nitrogen gas through a condenser to remove HCl, and was then trapped as zinc sulfide in a 25% zinc acetate solution. To moderate potential artifacts of hot distillation methods including elevated rates in control samples, experiments were analyzed in triplicate, on different days and with different glassware to minimize cross-contamination, and any activity observed in “control” samples was deleted from the treatments. The radioactivity of the resulting sulfide (Zn35S) and the remaining sulfate from the supernatant (35SO42-) were measured via liquid scintillation counter in Ultima Gold scintillation cocktail (ThermoFisher Inc., Waltham, MA).
Once on board ship, tubeworms and other macrofauna were removed from the samples and the large pieces were broken into more manageable fragments (~10-20 cm3) with a flame-sterilized chisel and sledgehammer, with the user wearing sterile nitrile gloves. Samples were quickly transferred to 0.2 um-filtered anaerobic (nitrogen-sparged) seawater. Samples were further broken down into smaller sizes while in anaerobic water, and subsamples from the interior of the fragments were immediately transferred to gastight jars (Freund Container Inc.) filled with sterile anaerobic seawater containing 2 mM sodium sulfide at pH 6, and stored at 4 degrees celsius for incubations and analyses. The sterile sulfidic seawater in the gastight jars were refreshed periodically during storage at 4 degrees celsius. The majority of the rate experiments (80%) were set up immediately on the ship using freshly collected samples. In parallel, subsamples (~1 cm3) from each flange were preserved aboard ship in glutaraldehyde (2.5% in phosphate buffered saline, PBS, pH 7.0), then prepared for electron microscopy via ethanol dehydration and critical point drying before being sputtered with a thin layer of gold-palladium to improve image resolution. Samples were imaged with a Zeiss model EVO Scanning Electron Microscope (SEM).
Prior to incubation, each flange subsample was pulverized by hand for about one hour to minimize fine-scale geological and microbial heterogeneity and facilitate more accurate experimental replication (akin to slurry experiments in sediments; Fossing & Jørgensen 1989; Weber & Jørgensen 2002; Jørgensen et al. 1992). Specifically, each subsample was pulverized with a flame-sterilized sledgehammer in sterile seawater actively bubbled with nitrogen within an anaerobic chamber. For each independent treatment, aliquots of 7.5 mL flange slurry (approx. 29 g wet weight and 20 g dry weight) were transferred into Balch tubes in an anaerobic chamber, and supplemented with 15 mL of sterile artificial seawater media designed to mimic the geochemical conditions within a hydrothermal flange (400 mM NaCl, 25 mM KCl, 30 mM CaCl2, 2.3 mM NaHCO3, 14 mM NaSO42-, 1 mM H2S, and 50 uM dissolved organic carbon – consisting of equimolar proportions 10 uM of pyruvate, citrate, formate, acetate, lactate) under a pure nitrogen headspace.
Concentrations of sulfide, sulfate and dissolved organic carbon (DOC) were varied independently to investigate concentration dependent effects on the rates of SR. The range of experimental conditions tested was determined from previously published concentration profiles of aqueous species modeled as functions of temperature and position within the Grotto vent structure (Tivey, 2004). Concentrations were varied by orders of magnitude within the modeled ranges to simulate conditions representative of different mixing regimes between seawater and vent fluid (Table 1). The range of DOC (which we approximate as a mix of pyruvate, citrate, formate, acetate, lactate – most of which have been identified to varying degrees within vent fluid and are known carbon sources for heterotrophic SR in culture) concentrations tested were based on the average DOC concentrations measured within diffuse fluids at the Main Endeavor Field (Lang et al., 2006; Lang et al., 2010). Hydrogen sulfide was present as H2S (pKa in seawater of 6.60) across all the conditions tested (Amend & Shock, 2001). Incubations were carried out at pH 4 (to simulate the pH of end-member Grotto vent fluid and the average calculated pH of mixed fluids in highly reduced zones within the flange; Tivey 2004) as well as pH 6 (representative of the calculated pH in fluid mixing zones; Tivey 2004). All the results are presented and discussed in the context of the initial measured media conditions.
Sufficient 35SO42- was added to achieve 15 uCi of activity. Samples were incubated anaerobically for 1, 3 or 7 days at ambient seawater (4 degrees celsius), thermophilic (50 degrees celsius) and hyperthermophilic (90 degrees celsius) temperatures. The range of temperatures considered was representative of different thermal regimes associated with the surface, outer layer and middle regions of hydrothermal chimneys (Tivey 2004; Kormas et al. 2006; Schrenk et al. 2003). Negative controls consisted of samples amended with 28 mM molybdate to inhibit SR (Newport & Nedwell, 1988; Saleh et al., 1964). Three biological replicates were run for each treatment, and two biological replicates for each control.
Upon completion, reactions were quenched with the injection of 5 mL 25% zinc acetate, at pH 8 (i.e. 20-fold excess Zn), and all samples were frozen at -20 degrees celsius for further analysis. 80% of incubations were performed shipboard with freshly collected samples and the remaining 20% of incubations were completed within one year of collection.
To determine SR rates, samples were thawed and the supernatant was removed and filtered through a 0.2 um syringe filter. The homogenized flange that remained in the tube was washed three times with deionized water to remove any remaining sulfate. One gram (wet weight) of flange material was added to 10 mL of a 1:1 ethanol to water solution in the chromium distillation apparatus, and then degassed with nitrogen for 15 minutes to drive the environment anoxic. Hydrogen sulfide gas was evolved after the anaerobic addition of 8 mL of 12 N HCl and 10 mL of 1 M reduced chromium chloride, followed by 3 hours of heating. The resulting hydrogen sulfide gas was carried via nitrogen gas through a condenser to remove HCl, and was then trapped as zinc sulfide in a 25% zinc acetate solution. To moderate potential artifacts of hot distillation methods including elevated rates in control samples, experiments were analyzed in triplicate, on different days and with different glassware to minimize cross-contamination, and any activity observed in “control” samples was deleted from the treatments. The radioactivity of the resulting sulfide (Zn35S) and the remaining sulfate from the supernatant (35SO42-) were measured via liquid scintillation counter in Ultima Gold scintillation cocktail (ThermoFisher Inc., Waltham, MA).
Rates were determined using the following calculation as in (Fossing & Jorgensen, 1989).

Where nSO42- is the quantity (in moles) of sulfate added to each incubation (14 mM * 15 mL = 210 umol), a is the activity (dpm) of the trapped sulfide, 1.06 is the fractionation factor between the hydrogen sulfide and sulfate pools (Jørgensen & Fenchel 1974), A is the activity of the sulfate pool at the completion of the incubation and t is the incubation time (days). The rates are presented in units of nmol S g-1 day-1. As previously mentioned, SR rates are numerically presented as the difference in rates between experimental and the molybdate inhibited controls, further mitigating any potential artifacts caused by hot distillation methods.
BCO-DMO Data Processing Notes:
-reformatted column names to comply with BCO-DMO standards
-filled in all blank cells with nd
-removed spaces and replaced with underscores
Tubeworm and macrofauna subsamples were imaged.
Instruments that generate enlarged images of samples using the phenomena of reflection and absorption of electrons behaving as waves.
pH sensor
DOC was measured
Used aboard ship and in lab
A device on shipboard or in the laboratory that holds water samples under controlled conditions of temperature and possibly illumination.
Used to quantify activity
concentration of sulfide
Concentration of sulfate (SO4) per unit volume
pH: The measure of the acidity or basicity of an aqueous solution
Temperatures at which samples were incubated anaerobically for 1, 3, or 7 days. 4 C: ambient seawater; 50 C: thermophilic; 90 C: hyperthermophilic.
Temperature in degrees C of a sample or other item. A generic temperature measurement.
Note: This is NOT water temp or sea surface temp
Duration of sample incubation; used in laboratory experiments. Refer to dataset for units of measure.
Concentration of sulfate (SO4) per unit volume
Concentration of sulfate (SO4) per unit volume
Concentration of sulfate (SO4) per unit volume
Latitude
latitude, in decimal degrees, North is positive, negative denotes South; Reported in some datasets as degrees, minutes
| Name | Affiliation | Contact |
|---|---|---|
| Peter R. Girguis | Harvard University | |
| Kiana L. Frank | Harvard University | |
| Hannah Ake | University of Hawaii at Manoa (SOEST) | ✓ |
| Shannon Rauch | University of Hawaii at Manoa (SOEST) | ✓ |
| Project Title | Characterizing the distribution and rates of microbial sulfate reduction at Middle Valley hydrothermal vents |
|---|---|
| Acronym | Middle Valley Vents |
| URL | https://www.bco-dmo.org/project/626603 |
| Created | November 17, 2015 |
| Modified | November 19, 2015 |
This project characterizes rates of microbially mediated sulfate reduction from three distinct hydrothermal vents in the Middle Valley vent field along the Juan de Fuca Ridge, as well as assessments of bacterial and archaeal diversity, estimates of total biomass and the abundance of functional genes related to sulfate reduction, and in situ geochemistry. Maximum rates of sulfate reduction occurred at 90°C in all three deposits. Pyrosequencing and functional gene abundance data reveal differences in both biomass and community composition among sites, including differences in the abundance of known sulfate reducing bacteria. The abundance of sequences for Thermodesulfovibro-like organisms and higher sulfate reduction rates at elevated temperatures, suggests that Thermodesulfovibro-like organisms may play a role in sulfate reduction in warmer environments. The rates of sulfate reduction observed suggest that – within anaerobic niches of hydrothermal deposits – heterotrophic sulfate reduction may be quite common and might contribute substantially to secondary productivity, underscoring the potential role of this process in both sulfur and carbon cycling at vents.
This project was funded, in part, by a C-DEBI Graduate Student Fellowship.
| Name | Affiliation | Role |
|---|---|---|
| Peter R. Girguis | Harvard University | Principal Investigator |
| Kiana L. Frank | University of Hawaii at Manoa (SOEST) | Contact |
| URL | https://www.bco-dmo.org/dataset/650324 |
|---|---|
| Download URL | https://www.bco-dmo.org/dataset/650324/data/download |
| Media Type | text/tab-separated-values |
| Created | June 29, 2016 |
| Modified | June 6, 2019 |
| State | Final no updates expected |
| Brief Description | O2 consumption of two strains of Mariprofundus ferrooxydans at two O2 conc. |
The cultivars of the neutrophilic iron oxidizers Mariprofundus ferrooxydans PV-1 and “TAG-1” were obtained from the Emerson lab at Bigelow laboratories.
These data were collected by placing each strain in a 100 mL serum vial with 6 mL of their standard, published media. The headspace was filled with the appropriate oxygen concentration by using bottled gas mixes and a regulator to flush the headspace without overpressurization. Prior to sealing the serum vials, a Presens OPTODE dot (sensor) was placed inside the vial, allowing non-invasive gas sampling of the changes in O2 in the headspace. A Presens four channel system was used to measure changes in oxygen concentration in realtime in each bottle. A total of four channels were measured during each experiment: channels 1 through 3 are the biological treatments and channel 4 was a kill control (microbe were killed through 24 hours of exposure to gamma radiation).
DMO notes:
Made top level .dat file from two elements in the filename: strain and O2 concentration
Removed units from data set, including umols/L and degrees C
changed treatment 4 to kill
changed from channel to treatment for column name
normalized variable names to BCO-DMO standards
A Presens™ OPTODE dot (sensor) was placed inside the sample vial, allowing non-invasive gas sampling of the changes in O2 in the headspace.
For more information about the PreSens OPTODE sensor, read this.
brief description, open ended, specific to the data set in which it appears
Oxygen; dissolved; reported in units of micromoles/liter
Experimental conditions applied to experimental units. In comparative experiments, members of the complementary group, the control group, receive either no treatment or a standard treatment.
local month, day and year, usually as a text string, e.g. feb10_1995. It is better to use one of the other forms of presenting date and time data so that the data can be used in computations and for comparisons. Note, if the string begins with numbers but also includes letters, there may be problems using the field name for retrieval.
time of day, local time, using 2400 clock format
Oxygen; dissolved; reported in units of micromoles/liter
date and time formatted to ISO 8601 standard; added by DMO. in the format YYYY-mm-ddTHH:MM:SS.xx.
Date/Time (Local) ISO formatted
This standard is based on ISO 8601:2004(E) and takes on any of the following forms:
YYYY-MM-DDTHH:MM:SS[.xx]
2009-08-30T09:05:00[.xx] (local time)
2009-08-30T14:05:00[.xx]Z (UTC time)
2009-08-30T14:05:00[.xx]-05:00
The dashes and the colons can be dropped.
The T can also be dropped "by mutual agreement", but one needs the trailing Z if the time is UTC.
Note the Time Zone (TZ) as +/-HH:MM
Time Zone signage is for conversion from local to UTC
West Coast USA in summer is +7 (add 7 hrs to local time for UTC)
| Name | Affiliation | Contact |
|---|---|---|
| Peter R. Girguis | Harvard University | |
| David Emerson | Harvard University | |
| David Johnston | Bigelow Laboratory for Ocean Sciences | |
| Jacob Cohen | Bigelow Laboratory for Ocean Sciences | |
| Hannah Ake | Harvard University | |
| Hannah Ake | Harvard University | |
| Hannah Ake | Woods Hole Oceanographic Institution (WHOI BCO-DMO) |
| Project Title | Collaborative Research: The Role of Iron-oxidizing Bacteria in the Sedimentary Iron Cycle: Ecological, Physiological and Biogeochemical Implications |
|---|---|
| Acronym | SedimentaryIronCycle |
| URL | https://www.bco-dmo.org/project/544584 |
| Created | January 8, 2015 |
| Modified | June 1, 2018 |
Iron is a critical element for life that serves as an essential trace element for eukaryotic organisms. It is also able to support the growth of a cohort of microbes that can either gain energy for growth via oxidation of ferrous (Fe(II)) to ferric (Fe(III)) iron, or by utilizing Fe(III) for anaerobic respiration coupled to oxidation of simple organic matter or H2. This coupled process is referred to as the microbial iron cycle. One of the primary sources of iron to the ocean comes from dissolved iron (dFe) that is produced through oxidation and reduction processes in the sediment where iron is abundant. The dFe is transported into the overlaying water where it is an essential nutrient for phytoplankton responsible for primary production in the world’s oceans. In fact, iron limitation significantly impacts production in as much as a third of the world’s open oceans. The basic geochemistry of this process is understood; however important gaps exist in our knowledge about the details of how the iron cycle works, and how critical a role bacteria play in it.
Intellectual Merit. Conventional wisdom holds that most of the iron oxidation in sediments is abiological, as a result of the rapid kinetics of chemical iron oxidation in the presence of oxygen. This proposal aims to question this conventional view and enhance our understanding of the microbes involved in the sedimentary iron cycle, with an emphasis on the bacteria that catalyze the oxidation of iron. These Fe-oxidizing bacteria (FeOB) utilize iron as a sole energy source for growth, and are autotrophic. They were only discovered in the ocean about forty-five years ago, and are now known to be abundant at hydrothermal vents that emanate ferrous-rich fluids. More recently, the first evidence was published that they could inhabit coastal sediments, albeit at reduced numbers, and even be abundant in some continental shelf sediments. These habitats are far removed from hydrothermal vents, and reveal the sediments may be an important habitat for FeOB that live on ferrous iron generated in the sediment. This begs the question: are FeOB playing an important role in the oxidative part of the sedimentary Fe-cycle? One important attribute of FeOB is their ability to grow at very low levels of O2, an essential strategy for them to outcompete chemical iron oxidation. How low a level of O2 can sustain them, and how this might affect their distribution in sediments is unknown. In part, this is due to the technical challenges of measuring O2 concentrations and dynamics at very low levels; yet these concentrations could be where FeOB flourish. The central hypothesis of this proposal is that FeOB are more common in marine sedimentary environments than previously recognized, and play a substantive role in governing the iron flux from the sediments into the water column by constraining the release of dFe from sediments. A set of experimental objectives are proposed to test this. A survey of near shore regions in the Gulf of Maine, and a transect along the Monterey Canyon off the coast of California will obtain cores of sedimentary muds and look at the vertical distribution of FeOB and putative Fe-reducing bacteria using sensitive techniques to detect their presence and relative abundance. Some of these same sediments will be used in a novel reactor system that will allow for precise control of O2 levels and iron concentration to measure the dynamics of the iron cycle under different oxygen regimens. Finally pure cultures of FeOB with different O2 affinities will be tested in a bioreactor coupled to a highly sensitive mass spectrometer to determine the lower limits of O2 utilization for different FeOB growing on iron, thus providing mechanistic insight into their activity and distribution in low oxygen environments.
Broader Impacts. An important impact of climate change on marine environments is a predicted increase in low O2 or hypoxic zones in the ocean. Hypoxia in association with marine sediments will have a profound influence on the sedimentary iron cycle, and is likely to lead to greater inputs of dFe into the ocean. In the longer term, this increase in dFe flux could alleviate iron-limitation in some regions of the ocean, thereby enhancing the rate of CO2-fixation and draw down of CO2 from the atmosphere. This is one important reason for developing a better understanding of microbial control of sedimentary iron cycle. This project will also provide training to a postdoctoral scientist, graduate students and undergraduates. This project will contribute to a student initiated exhibit, entitled ‘Iron and the evolution of life on Earth’ at the Harvard Museum of Natural History providing a unique opportunity for undergraduate training and outreach.
| Name | Affiliation | Role |
|---|---|---|
| David Emerson | Bigelow Laboratory for Ocean Sciences | Principal Investigator |
| Peter R. Girguis | Harvard University | Principal Investigator |
| David Johnson | Harvard University | Co-Principal Investigator |